Регуляція експресії генів в еукаріотичних клітинах
- 30-07-2021, 22:09
- 1 313
10 Клас , Біологія і екологія 10 клас Остапченко (рівень стандарту)
§ 29. РЕГУЛЯЦІЯ ЕКСПРЕСІЇ ГЕНІВ В ЕУКАРІОТИЧНИХ КЛІТИНАХ
Пригадайте будову еукаріотичних генів і геномів, етапи і механізми реалізації генетичної інформації. Що таке хроматин і в яких станах він може перебувати? Які функції РНК-полімерази? Який вплив гормонів на організм?
Активність генів в еукаріотичних клітинах. У клітинах еукаріотичних організмів кількість структурних (білкових) генів може досягати декількох десятків тисяч. Зрозуміло, що одночасна робота всіх цих генів не потрібна, оскільки, наприклад, для будь-якої клітини було б марнотратством продукувати ферменти, які в даний час непотрібні для синтезу продуктів, що і так є в достатній кількості. Щоб зберегти енергію, запас органічних і мінеральних сполук, клітини регулюють активність своїх генів так, що в даний момент синтезуються продукти лише тих генів і в тих кількостях, які потрібні для нормального функціонування клітини.
В еукаріотичних клітинах активні приблизно 15 % усіх генів. У багатоклітинних організмів у різних типах клітин експресуються різні набори генів: гени, активні в клітинах одного типу, можуть бути неактивними в клітинах іншого. Це є наслідком диференціації клітин в онтогенезі. Саме активність відповідних генів визначає характерні риси і роль, яку певні клітини виконують в організмі.
Експресія генів регулюється на будь-якому етапі реалізації генетичної інформації: на рівні транскрипції відповідного гена, дозрівання мРНК, регуляції тривалості життя мРНК, на рівні трансляції та функціонування кінцевого продукту в результаті його модифікацій. На сьогодні найкраще вивчено механізми регуляції транскрипції. Є два основні механізми, які дають змогу контролювати ефективність цього процесу: використання додаткових білкових факторів транскрипції та зміни упакування хроматину.
Фактори транскрипції (транскрипційні фактори) — білкові молекули, які контролюють процес транскрипції (синтезу мРНК на матриці ДНК), зв’язуючись зі специфічними ділянками молекули ДНК. Свої функції активатора або пригнічувача (репресора) вони виконують самостійно або в комплексі з іншими білками.
Регуляція експресії генів за допомогою білкових факторів. Додаткові транскрипційні фактори — це велика родина білків, які регулюють експресію генів, зв’язуючись із короткими регуляторними послідовностями молекул ДНК (мал. 29.1). Додаткові транскрипційні фактори впізнають регуляторні послідовності ДНК, розташовані або близько від промотора, або на відстані кількох тисяч нуклеотидів від гена, активність якого регулюється. Вони можуть або активувати, або знижувати експресію цього гена. Точність регуляції транскрипції відповідних генів зумовлена тим, що кожний специфічний транскрипційний фактор упізнає тільки власну ДНК-послідовність.

Мал. 29.1. Транскрипційні фактори впізнають регуляторні елементи і, взаємодіючи з РНК-полімеразою, підсилюють чи послаблюють роботу гена
Експресія конкретного гена залежить не від одного регуляторного білка, а від їхніх комплексів, тому існує достатньо великий набір комбінацій, яких вистачає для регуляції транскрипції всіх потрібних у певний момент для клітини генів. Тому один і той самий білок може бути і активатором, і репресором.
Запам'ятаємо
Гени, які кодують білки, що регулюють активність інших генів, називають генами-регуляторами.
Часто клітина повинна оперативно реагувати на зміни навколишнього середовища (почати поділ, синтез потрібних речовин тощо), тобто швидко запустити роботу потрібних генів. Для цього вона вже має деяку кількість потрібних регуляторних білків, що перебувають у неактивному стані. Їхня активація залежить від впливу зовнішніх специфічних сигнальних молекул. Роль зовнішніх сигналів, які змінюють експресію генів у клітині, відіграють гормони.
Активація наявних регуляторних білків часто відбувається за рахунок утворення комплексу з відповідними молекулами-акцепторами або в результаті хімічних модифікацій амінокислот. У результаті цих процесів білок змінює свою просторову структуру і переходить з неактивного стану в активний (мал. 29.2).
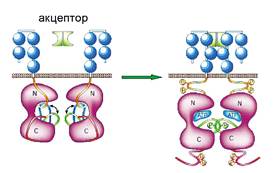
Мал. 29.2. Активація рецептора під час зв’язування з молекулою-акцептором
Так, сигнальні молекули зв’язуються з рецепторами на поверхні клітинної мембрани клітини-мішені й запускають усередині клітини каскад реакцій, який забезпечує передачу сигналу від клітинної мембрани до ядра (мал. 29.3). Саме за таким типом передаються у клітину сигнали, що спонукають її до мітотичного поділу.

Мал. 29.3. Схема активації роботи генів, які регулюються стероїдними або тиреоїдними гормонами: гормон проходить крізь мембрану клітини (а), зв’язується з внутрішньоклітинним рецептором (б), який транспортується в ядро, де активує гени-мішені (в)
Вплив упакування хроматину на ефективність транскрипції еукаріотичних генів. Ви вже знаєте, що активний синтез молекул РНК відбувається в неущільнених ділянках хроматину. Механізми активації чи пригнічення роботи еукаріотичних генів мають бути тісно пов’язані з процесами, що забезпечують конденсацію чи деконденсацію хроматину. Процеси зміни упакування хроматину достатньо складні та пов’язані з модифікаціями як основних білків хроматину (гістонів), так і ДНК.
Гістони у транскрипційно неактивному хроматині є метильованими, а в активному вони ацетильовані. Модифікації ДНК стосуються метилування амінокислоти цитозину, що займає певне положення в регуляторних ділянках деяких генів (мал. 29.4).

Мал. 29.4. Хімічні модифікації гістонів (червоним позначено залишки етанової (оцтової) кислоти, зеленим — метильні групи) та ДНК у транскрипційно активному й неактивному хроматині
Метилування — введення до складу органічної сполуки метильної групи (—СН3) замість атому Гідрогену (або металу чи галогену). Ацетилування — заміщення в молекулах органічних сполук атомів Гідрогену залишком етанової (оцтової) кислоти (ацетильною групою —CH3CO).
Під час активації експресії гена транскрипційні фактори взаємодіють з ферментами: одні з них здійснюють деметилування гістонів (процес протилежний метилуванню), інші — їх ацетилування, що приводить до декомпактизації хроматину. Під час пригнічення активності генів залучаються ферменти, які здійснюють деацетилування (процес, протилежний ацетилуванню) та метилування гістонів (у деяких випадках — метилування ДНК). Метильні групи гістонів є сигналами для білків, які збезпечують подальшу конденсацію хроматину (перехід його у стан гетерохроматину). Інколи цей процес буває необоротним і гени вимикаються назавжди.
Контроль експресії генів під час дозрівання мРНК. Крім описаних процесів регуляції транскрипції, в еукаріотів діють механізми, які регулюють експресію генів за рахунок процесів альтернативного дозрівання мРНК.
Альтернативне дозрівання мРНК — способи дозрівання мРНК, за яких з однієї послідовності гена виникають різні (хоча і подібні) послідовності мРНК. Воно може відбуватися в результаті двох процесів: альтернативного сплайсингу пре-мРНК та редагування мРНК. Під час альтернативного сплайсингу деякі екзони можуть бути видалені подібно до інтронів. У результаті може утворитися декілька альтернативних молекул мРНК. Альтернативний сплайсинг відповідної молекули мРНК може відбуватися в клітинах різного типу або в клітинах одного типу, але на різних етапах її розвитку.
У разі редагування мРНК послідовності мРНК змінюються в результаті вставляння або видалення нуклеотидів чи заміни нітрогеновмісних основ усередині нуклеотидів.
Регуляція експресії генів на рівні ефективності перебігу трансляції. Продуктивність трансляції може контролюватися специфічними білками, які в цитоплазмі зв’язуються з мРНК, утворюючи нуклеопротеїнові комплекси — інформосоми. Білки інформосом, з одного боку, захищають мРНК від розкладання на нуклеотиди, а з іншого — повністю блокують трансляцію. Отже, в клітині є «запас» довгоіснуючих молекул мРНК, які за потреби можуть бути використані. Інші білки, зв’язуючись з мРНК, можуть прискорювати розкладання мРНК або блокувати посадження рибосоми на молекулу мРНК.
Поширеним механізмом регуляції трансляції шляхом впливу на тривалість життя молекул мРНК є РНК-інтерференція. Якщо відбувається надлишкова транскрипція якогось гена, то починається синтез комплементарного ланцюга РНК на молекулі мРНК. Двониткова РНК розпізнається специфічним ферментом і нарізається на невеликі двониткові фрагменти з 20—30 нуклеотидних пар. Після денатурації фрагментів нитки комплементарного ланцюга РНК зв’язуються з мРНК і запускають трансляцію наново (мал. 29.5).
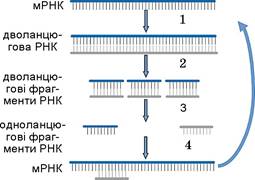
Мал. 29.5. Схема мРНК-інтерференції: 1 — синтез комплементарного ланцюга РНК; 2 — розрізання дволанцюгової РНК на фрагменти; 3 — денатурація фрагментів; 4 — приєднання антисенсорного фрагмента до мРНК
Ключові терміни та поняття
фактори транскрипції, гени-регулятори, альтернативне дозрівання мРНК, РНК-інтерференція.
Перевірте здобуті знання
1. У чому полягає необхідність регуляції активності генів у клітині? 2. Чим можна пояснити те, що в клітинах різних типів одного організму активні різні групи генів? 3. Як відбувається регуляція експресії генів за допомогою білкових факторів? 4. Що таке гени-регулятори? 5. Як гормони різної біохімічної природи впливають на експресію генів? 6. Як здійснюється контроль за експресією генів під час дозрівання мРНК?
Поміркуйте
У чому полягає біологічне значення явища РНК-інтерференції?
Задача для самостійного розвитку
Розрахуйте кількість генів, яку може регулювати комплекс із 10 транскрипційних факторів.
Коментарі (0)