Основні закономірності функціонування генів у еукаріотів
- 12-05-2021, 10:19
- 996
11 Клас , Біологія і екологія 11 клас Балан, Вервес (рівень стандарту, академічний рівень)
§ 14. Основні закономірності функціонування генів у еукаріотів
Аби краще засвоїти матеріал цього параграфа, слід пригадати: що таке ген, геном, регуляторні та структурні гени, екзони, інтрони, сплайсинг? Як відбувається синтез білкових молекул? Які з гормонів людини мають стероїдну та білкову природу? Які існують механізми регуляції активності генів у прокаріотів? Яка будова хромосоми? Яка будова гемоглобіну?
Які особливості організації геному еукаріотів?
В еукаріотів значна частина геному представлена послідовностями нуклеотидів, які не кодують структури молекул білків та РНК. Учені з’ясували, що 20-50 % генів еукаріотів мають одну, інші - кілька зазвичай не ідентичних копій. Понад 50 % усієї кількості ДНК припадає на повтори. Завдяки копіям, мутації окремих генів мало впливають на фенотип. Повторювані послідовності нуклеотидів зосереджені переважно на кінцях хромосом і в зоні прикріплення ниток веретена поділу (центромера).
Генам еукаріотів, на відміну від генів прокаріотів, притаманний мозаїчний характер будови: кодуючі ділянки (екзони) чергуються з некодуючими (інтронами). Серед інтронів є ділянки, що здійснюють важливі регуляторні функції. Регуляторні ділянки є й у складі міжгенної ДНК. Обов’язковим етапом, необхідним для здійснення молекулою іРНК своїх функцій, є процес сплайсингу: інтрони за участі відповідних ферментів вирізаються, а екзони зшиваються, утворюючи матрицю для синтезу білкової молекули (див. мал. 9.1).
Які особливості регуляції експресії генів у еукаріотів?
На відміну від прокаріотів, у клітинах еукаріотів молекули іРНК синтезуються на молекулах ДНК в ядрі і вже звідти транспортуються до цитоплазми. Таким чином, такі основні етапи експресії гена, як транскрипція та трансляція (синтез білкової молекули на матриці іРНК) в еукаріотів розділені у просторі та часі.
Гени багатоклітинних організмів поділяють на дві функціональні групи: одні з них активні в усіх клітинах, де забезпечують універсальні функції живого, а інші - активні лише у клітинах певних типів. Особливу групу становлять псевдогени - змінені внаслідок мутацій послідовності нуклеотидів, які нагадують певні гени, однак інформація з них не зчитується.
До регуляторних елементів експресії генів належать промотори (послідовність нуклеотидів, з якої розпочинається процес транскрипції), послідовності нуклеотидів, які впливають на ефективність процесу транскрипції, а також ті, що забезпечують припинення транскрипції та трансляції.
Запам’ятайте: на активність генів еукаріотів і прокаріотів впливають певні регуляторні послідовності нуклеотидів, які функціонально пов’язані з ними.
Гени еукаріотів, які керують процесами транскрипції, не можуть бути постійно активними. Тому гени та їхні функціональні групи під впливом певних чинників чи фаз клітинного циклу то активуються, то через певний час знову стають неактивними. Завдяки тому, що не всі гени клітини активні одночасно, різні типи клітин продукують не весь закодований комплекс білків чи молекул РНК, а лише потрібні на даний момент. Так, гени, які кодують структуру субодиниць гемоглобіну дорослої людини, активні лише у клітинах-попередниках еритроцитів. Здатність клітин «вмикати» або «вимикати» структурні гени має важливе значення і для економного витрачання енергетичних ресурсів.
Запам’ятайте: активність будь-якого структурного гена залежить від наявності в клітині чинників, які пригнічують процеси транскрипції або стимулюють їх.
Для регуляції активності певних генів використовується механізм, за якого особливі білкові молекули запобігають розпізнаванню ферментом, що забезпечує синтез іРНК, сигналу про припинення синтезу цієї молекули. Такий сигнал становить певну послідовність нуклеотидів, розташовану всередині кодуючої частини гена (пригадайте, які саме триплети сигналізують про припинення синтезу білкової молекули). За наявності факторів, які запобігають розпізнаванню ферментом сигналу про припинення синтезу іРНК, синтез цієї молекули продовжується і вона стає функціонально активною.
Деякі неактивні гени можуть активуватись позаклітинним сигналом, наприклад впливом підвищеної температури чи певних гормонів. Так, потрапивши до клітини, стероїдний гормон взаємодіє з клітинним білком і змінює його структуру. Змінений білок проникає у ядро і зв’язується зі специфічним регуляторним елементом, який ініціює транскрипцію відповідного гена - синтез молекули іРНК.
Гормони білкової природи зв’язуються з відповідним рецептором, розташованим на зовнішньому боці мембрани. Це активує фермент, за участі якого починає діяти фактор транскрипції певного гена.
Деякі білкові молекули здатні переміщуватись у внутрішньому середовищі клітини, «шукаючи» відповідні регуляторні послідовності нуклеотидів. Їхня взаємодія може сприяти активації транскрипції або блокувати цей процес. Перехід певних генів в активний чи неактивний стан можливий і при взаємодії цих білкових молекул з ферментами, що каталізують синтез іРНК.
Запам’ятайте: існують послідовності нуклеотидів, що межують, перекриваються або розташовані поза промотором, з якими взаємодіють регуляторні білки. Ці білки можуть бути активаторами або пригнічуваними (дипресорами), а також запобігати завершенню транскрипції.
Як ви пам’ятаєте, структурною частиною хромосоми є нуклеосома (мал. 14.1). Вона формується за рахунок взаємодії нитки ДНК з білками (гістонами). Нуклеосоми є загальними пригнічувачами активності генів. Більшість генів у клітині перебуває в неактивному стані, за винятком активованих факторами транскрипції. Активація транскрипції потребує перебудови хроматинових ниток. Ця перебудова пов’язана з тим, що певні ділянки хроматинових ниток стають менш ущільненими і регуляторні ділянки молекули ДНК, пов’язані з даним геном, звільняються від нуклеосом. Так відповідні ділянки ДНК стають доступними для чинників, які забезпечують процес транскрипції.
На експресію генів можуть впливати й зміни хімічного складу ДНК, але послідовність нуклеотидів у молекулі при цьому не змінюється. Так, у певних ділянках може відбуватись введення метильної групи (-СН3) (процес метилювання). Така модифікація молекули ДНК не супроводжується зміною нуклеотидної послідовності. Підвищення ступеня метилювання у певних ділянках спричиняє зменшення або повне пригнічення експресії сусідніх генів. І навпаки, зменшення ступеня метилювання пов’язане з підвищенням рівня їхньої експресії.
Під час формування специфічної природної структури ДНК регуляторні послідовності і відповідні структурні гени зближуються, що полегшує їхню взаємодію. Крім того, фактори транскрипції (наприклад, певні білки) зв’язуються з регуляторними послідовностями нуклеотидів і можуть утворювати ланцюжок, який зближує віддалені структурні гени та їхні регуляторні ділянки.
Активність генів може залежати від взаємодії молекул нуклеїнових кислот між собою. Одним з таких механізмів є РНК-інтерференція (від лат. інтер - між, посеред і ферентіс - який несе) - процес пригнічення експресії гена за допомогою молекул мікро-РНК. Це пригнічення експресії гена відбувається під час транскрипції, трансляції або руйнування іРНК.
Молекули мікро-РНК не кодують структуру білків і мають незначну довжину: 19-21 пара нуклеотидів. Як вони виникають? Активація певних генів зумовлює синтез молекул РНК з дволанцюговими ділянками. Ці ділянки вирізаються за участі особливого ферменту, З них утворюються молекули мікро-РНК, кожна з яких зв’язується з кількома білковими молекулами, що забезпечують розплітання ланцюгів РНК. Згодом один з ланцюгів мікро-РНК сполучається з комплементарною ділянкою іРНК, яка кодує структуру певного гена. Наслідки такої взаємодії бувають різні. По-перше, це може бути ущільнення хроматину та пригнічення активності гена. По-друге, після транскрипції молекула іРНК може зруйнуватись, що унеможливлює синтез білка (переважно в рослинних клітинах). По-третє, можлива зупинка синтезу білка (переважно в тваринних клітинах). Є дані, що молекули мікро-РНК можуть також безпосередньо взаємодіяти з молекулами ДНК.
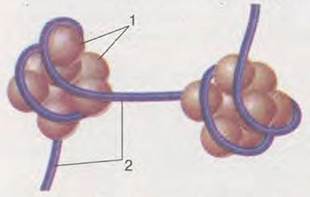
Мал. 14.1. Структура нуклеосоми: 1 - глобулярні білки; 2 - нитка ДНК
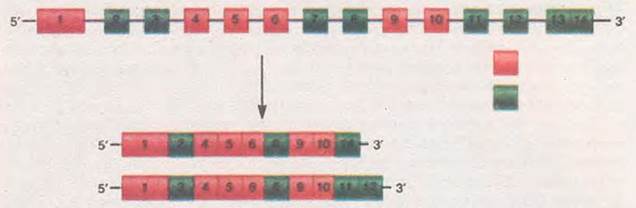
Мал. 14.2. Альтернативний сплайсинг: вгорі - молекула незрілої іРНК, яка кодує білок a-тропоміозин: червоним кольором показано екзони, які обов’язково залишаються у зрілій молекулі іРНК (1, 4-6, 9, 10), зеленим (2, 3, 7, 8, 11-14) - необов’язково. Зверніть увагу: залежно від того, які саме необов’язкові екзони залишаються після альтернативного сплайсингу, утворюються різні молекули іРНК: верхня кодує білок, що входить до складу непосмугованих м’язових клітин, нижня - посмугованих
Запам’ятайте: експресія генів може залежати від взаємодії молекул нуклеїнових кислот між собою або змін їхнього хімічного складу.
Ще один шлях регуляції експресії генів - це альтернативний сплайсинг. Ми згадували, що сплайсинг - процес, пов’язаний з видаленням з іРНК ділянок, що відповідають інтронам. Але він може перебігати й альтернативним1 шляхом (звідки й походить його назва). Наприклад, за участі певних ферментів можуть вирізатись декілька екзонів як на початку, так і всередині гена. У результаті цього з однієї ділянки можуть бути синтезовані різні варіанти молекул іРНК, які кодують різні білки (мал. 14.2). У геномі людини середня кількість альтернативних форм іРНК, які припадають на один ген, становить 5,4.
Запам’ятайте: експресія генів може пригнічуватися під час транскрипції або трансляції. При цьому послідовність генів може не змінюватись.
Як результати вивчення активності генів застосовують у генетичній інженерії?
Генетична інженерія - це експериментальна галузь молекулярної генетики. Її основні завдання:
- синтез поза організмом, виділення з клітин і перебудова окремих генів;
- копіювання і розмноження виділених або синтезованих генів;
- введення генів або їхніх груп до геномів інших організмів;
- поєднання різних геномів в одній клітині.
1 Альтернатива (від лат. альтернаре - чергуватися, вагатися, альтер - один з двох) - необхідність вибору між двома можливостями, що виключають одна одну.
Важливим етапом цих досліджень є встановлення нуклеотидних послідовностей окремих генів. Без цього неможливо виділяти окремі гени з організму або здійснювати їхній синтез поза організмом. Але недостатньо просто ввести певний ген у геном іншого організму. Вся ця робота втратить свій сенс, якщо не відбудеться експресії такого гена. А вона неможлива без урахування різноманітних чинників, які впливають на неї.
Регуляторні послідовності нуклеотидів еукаріотів за своєю структурою та розташуванням відносно кодону, який ініціює процес транскрипції, відрізняються від подібних послідовностей у клітинах прокаріотів. Якщо гени еукаріотів просто ввести у прокаріотичну клітину, бактеріальний фермент, який забезпечує синтез іРНК, не розпізнає ці послідовності. Тому для експресії генів еукаріотів у клітинах прокаріотів потрібно, щоб ці гени потрапили під контроль регуляторних елементів клітини хазяїна.
При перенесенні генів еукаріотів необхідно також враховувати, що в їхніх клітинах кожний структурний ген має власний набір регуляторних елементів, які активуються специфічними сигналами у різні періоди клітинного циклу. Натомість структурні гени прокаріотних клітин об’єднані у групи, що мають спільні елементи регуляції.
Досконале вивчення проблем експресії тих чи інших генів у клітині дало змогу вирішити деякі важливі практичні питання за допомогою генної інженерії. Ви знаєте, що відсутність або недостатність вироблення тих чи інших гормонів в організмі людини або тварин спричинює важкі наслідки. Зокрема, досить поширене захворювання людей - цукровий діабет - є наслідком втрати здатності до синтезу інсуліну; тому для підтримання життєдіяльності необхідно періодично вводити в організм цей гормон. Отримувати препарати інсуліну у достатній кількості дозволило введення відповідних генів у бактеріальну клітину.
РНК-інтерференцію можна використовувати у медичній практиці та селекції. Для цього створюють так звані антисенсорні (від грец. анти - протилежний та лат. сенсус - відчуття) молекули РНК. За складом вони відповідають послідовності нуклеотидів молекули іРНК певної клітини. Припустимо, що така сенсорна РНК несе інформацію про шкідливі для організму або небажані для селекціонера білки. Тоді вчені синтезують і вводять у клітину молекулу антисенсорної РНК. Її нуклеотиди зв’язуються з нуклеотидами сенсорної РНК. Це блокує біосинтез білків, закодованих у молекулі сенсорної РНК. Наприклад, застосовуючи антисенсорні технології, можна послабити симптоми ВІЛ-інфекції, коли антисенсорні молекули РНК зв’язують комплементарну іРНК вірусу, що гальмує його розмноження.
Застосування антисенсорної технології у селекції рослин має вже певні практичні результати. У помідорів відомі ферменти, які сприяють їхньому швидкому дозріванню. Але фермерам потрібно, щоб ці овочі достигали поступово, оскільки важко реалізувати значні обсяги продукції за короткий термін. Крім того, один з ферментів прискорює розщеплення полісахариду пектину в клітинних стінках, унаслідок чого плоди швидко псуються. Завдяки введенню у геном помідорів антисенсорних молекул РНК синтез відповідних ферментів гальмується.
Метод РНК-інтерференції застосовують для з’ясування деяких теоретичних питань при дослідженнях з використанням культур клітин або живих організмів. За рахунок синтезованих дволанцюгових молекул мікро РНК, які вибірково впливають па активність тих чи інших генів, можна з’ясувати їхні функції. При введенні молекули мікро-РНК вона пригнічує активність специфічного гена, і за наслідками цього процесу вчені роблять висновки щодо його функцій.
Нові терміни та поняття. Явище РНК-інтерференції, антисенсорні технології.
Запитання для повторення: 1. Чим відрізняються між собою організації геному еукаріотів і прокаріотів? 2. Яка роль сплайсингу в активації генів? 3. Як регулюється експресія генів за участю регуляторних білків? 4. Які відомі регуляторні послідовності нуклеотидів, що впливають на експресію генів? 5. У чому полягає явище РНК-інтерференції? 6. У чому полягає значення альтернативного сплайсингу?
Проблемне завдання. Поміркуйте, що спільного та відмінного між процесами транскрипції у вірусів та альтернативним сплайсингом в еукаріотів.
Коментарі (0)