Макроеволюційні процеси
- 16-05-2021, 13:31
- 985
11 Клас , Біологія і екологія 11 клас Балан, Вервес (рівень стандарту, академічний рівень)
§ 39. Макроеволюційні процеси
Аби краще засвоїти матеріал цього параграфа, слід пригадати: чи реально існують у природі надвидові систематичні групи (роди, родини тощо)? Навіщо їх використовують у науковій систематиці? Які є сучасні критерії виду? Що таке адаптивна радіація? Що вивчають палеонтологія та біогеографія?
Що таке макроеволюція?
Макроеволюція - сукупність еволюційних процесів, що приводять до виникнення надвидових таксонів (родів, родин і т. д. аж до царства).
Як відомо, реально в природі існують лише види, а надвидові систематичні категорії вчені ввели для їхнього впорядкування (систематизації): певні види належать до певного роду, певні роди - до певної родини тощо. Належність виду до того чи іншого роду, родів - до родини тощо дослідники встановлюють на основі ступеня їхньої історичної спорідненості. Для цього вони використовують різні критерії: морфологічні, генетичні, географічні тощо. Тому особливих механізмів макроеволюції не існує. Певні її закономірності є наслідком узагальнень ученими накопичених протягом історичного розвитку відмінностей між спорідненими видами, які виникають унаслідок тривалих мікроеволюційних змін і послідовного ряду видоутворень.

Мал. 39.1. Адаптивна радіація ссавців. Подумайте, які риси подібності виникли в різних груп унаслідок адаптацій до певного середовища життя (наземно-повітряного, водного, ґрунтового тощо)
Різноманітність видів зумовлена пристосуваннями організмів до різних умов довкілля, які супроводжуються дивергенцією. Це явище має назву адаптивна радіація. Наприклад, усі ряди плацентарних ссавців виникли від спільного предка внаслідок пристосувань до різних умов наземного, водного (ластоногі, китоподібні) та повітряного (рукокрилі) середовищ (мал. 39.1).
Що таке біологічні прогрес і регрес? Якими шляхами може бути досягнутий біологічний прогрес?
Вивчаючи історичний розвиток тварин, О.М. Сєверцов (мал. 37.1, 3) у 20-х роках XX сторіччя розробив учення про біологічні прогрес і регрес.
Біологічний прогрес проявляється в збільшенні чисельності популяцій, розширенні ареалу та утворенні нових підвидів і видів у межах певної групи. Наприклад, у стані біологічного прогресу в наш час перебувають покритонасінні, комахи, молюски, птахи, ссавці тощо. Він є наслідком еволюційного успіху певної групи.

Мал. 39.2. 1. Африканський слон. 2. Індійський слон. 3. Різноманіття викопних хоботних
Біологічний регрес - це наслідок неспроможності пристосуватись до змін довкілля. Він проявляється в зменшенні чисельності популяцій, звуженні ареалів та може призвести до вимирання певної групи. Наприклад, з колись багатого на види ряду хоботних нині залишились африканський та індійський слони, які перебувають під загрозою повного вимирання (мал. 39.2).
Неважко помітити, що, оскільки існують лише види, поняття про прогрес чи регрес стосується лише кожного з них окремо (мал. 39.3). За даними палеонтології, багато груп організмів безслідно зникли, однак це явище свідчить, насправді, лише про вимирання конкретних споріднених видів. З іншого боку, у кожній з «процвітаючих» груп багато видів зникають, їм на зміну виникають нові, які займають подібні екологічні ніші, однак зовсім не обов’язково є родичами зниклих. Наприклад, після вимирання динозаврів їхнє місце в наземних екосистемах зайняли ссавці. Цікаво, що динозаври - це лише два ряди плазунів, тоді як протягом сучасної (кайнозойської) ери повністю вимерли 14-16 рядів ссавців. Голонасінних нині налічують ледве 600 видів, а «примітивних» папоротей - 10 000!

Мал. 39.3. I. Види фауни та флори України, які перебувають у стані біологічного прогресу: 1 - сосна звичайна; 2 - хвощ польовий; 3 - метелик білан капустяний; 4 - карась сріблястий. II. Види фауни та флори України, які перебувають у стані біологічного регресу: 5 - едельвейс; 6 - тюльпан Шренка; 7 - жук-скарабей; 8 - поліксена
Уявлення про зміни будови організмів для досягнення біологічного прогресу мають узагальнюючий характер. До них належать поняття про ароморфоз, ідіоадаптацію та загальну дегенерацію (мал. 39.4). Ароморфоз - (від грец. айро - піднімаю та морфозис - форма, зразок), або морфофізіологічний прогрес, — еволюційне перетворення, яке підвищує рівень організації організму в цілому і відкриває нові можливості для пристосування до різноманітних умов існування. Наприклад, виникнення щелеп у хребетних дало їм можливість живитись великою здобиччю; утворення квітки у покритонасінних привело до урізноманітнення способів запилення та формування плодів тощо.
Загальна дегенерація (від лат. дегенеро - вироджуюсь), або морфофізіологічний регрес, - явище спрощення організмів у процесі еволюції. Відомо, що в паразитичних тварин зникають органи чуття, а також цілі системи органів (наприклад, травна - у стьожкових червів). Деякі паразитичні види рослин втрачають хлорофіл і відповідно - здатність до фотосинтезу (наприклад, петрів хрест; мал. 39.4, 1). Малорухомість і пасивний (фільтруючий) спосіб життя також ведуть до дегенерації - наприклад, у двостулкових молюсків, вусоногих рачків (мал. 39.4, 2) тощо. Інколи дегенерації зазнає лише певна фаза життєвого циклу (наприклад, гаметофіт у насінних рослин).
Результатом як ароморфозу, так і загальної дегенерації є розширення адаптаційних можливостей, яке реалізується за допомогою ідіоадаптацій. Ідіоадаптація (від грец. ідіос - особливий, своєрідний та лат. адаптаціо - пристосування) - певна зміна будови організму, яка має характер пристосування до певних умов та не змінює рівень його організації. Приклади ідіоадаптацій - це різна будова квіток покритонасінних (мал. 39.5), різноманітні ротові органи комах, різна форма дзьоба у птахів (мал. 34.7, 2), пристосована для здобування різної їжі та ін. Ідіоадаптації забезпечують адаптивну радіацію, однак організація всіх видів залишається на рівні предкової.

Мал. 39.4. Приклади загальної дегенерації: 1 - рослина петрів хрест паразитує на коренях дерев: її надземні органи позбавлені хлорофілу, але вона утворює пурпурові квітки, зібрані в китицю. Її багаторазово розгалужене кореневище розвиває тонке коріння, що присмоктується до коренів рослини-хазяїна; 2 - вусоногий рачок веде прикріплений спосіб життя; живиться за допомогою фільтрації води. Оточений карбонатним захисним будиночком; складні очі та черевце редукуються; кінцівки створюють потоки води і перетворені на органи фільтрації для вилучення з води твердих поживних частинок

Мал. 39.5. Різноманітність будови квіток покритонасінних
О.М. Сєверцов сформулював положення про співвідношення між собою основних шляхів досягнення біологічного прогресу (зміна фаз еволюційного процесу, або «закон Сєверцова»): як правило, у межах природної (тобто монофілетичної) групи організмів за періодом ароморфозів чи загальної дегенерації настає час ідіоадаптацій (мал. 39.6). Отже, під час історичного розвитку певної систематичної групи ароморфози чи загальна дегенерація відбуваються значно рідше, аніж ідіоадаптації.
Як здійснюється інтеграція живого на різних рівнях організації?
Різні рівні організації живої матерії співвідносяться так, що кожен вищий щабель містить у собі нижчий. Так, біомолекули складають клітини, клітини - організм, організми - популяції, а популяції, у свою чергу, - біоценози. Отже, довкілля впливає на нижчий рівень не безпосередньо, а через усі вищі від нього. Тому від факторів навколишнього середовища найбільше залежні біогеоценози, найменше - біомолекули, що зумовлює різні ступені інтеграції живого на кожному рівні його організації.
Інтеграція (від лат. інтегратіо - відновлення, поповнення) - структурне об’єднання та узгодження дій окремих частин, що входять до складу цілісної системи (наприклад, клітин в багатоклітинному організмі).
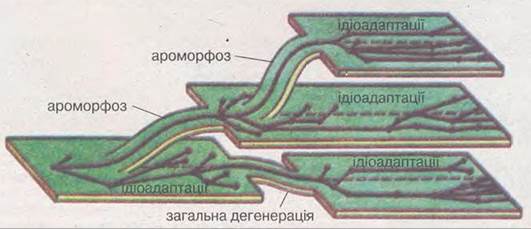
Мал. 39.6. Схема, що ілюструє співвідношення між ароморфозом, загальною дегенерацією та ідіоадаптаціями
Найвищий ступінь інтеграції здійснюється на молекулярному рівні, де величезна різноманітність молекул складає кілька стабільних типів сполук: білки, ліпіди, вуглеводи, нуклеїнові кислоти. Найнезначніші зміни структури молекул найважливіших біополімерів, зокрема білків і нуклеїнових кислот, можуть докорінно змінити їхні властивості. Така інтеграція - необхідна умова нормального функціонування надзвичайно складних молекулярних систем живих істот.
На клітинному рівні ступінь інтеграції зменшується: число та форма певних органел можуть значно змінюватись у клітині, не порушуючи її функцій тощо. На організмовому рівні інтеграція ще нижча: значно варіюють маса, ріст, забарвлення та багато інших ознак організму. Ще менш інтегровані популяції, у яких відбуваються постійні зміни числа особин, їхніх вікових груп, різних фаз розвитку тощо.
Інтегрованість у біогеоценозах дуже слабка і полягає лише в підтриманні колообігу речовин і перетворень енергії в ланцюгах живлення. Окремі ланки цих ланцюгів можуть легко взаємно замінюватись. Ступінь інтеграції зумовлює рівень ефективності використання енергії живою системою. Регуляторні механізми також значно більш жорсткі на нижчих рівнях порівняно з вищими.
Ще однією особливістю живих систем є їхня здатність функціонувати лише в стабільних умовах, тому на кожному рівні організації підтримується гомеостаз її внутрішнього середовища. Найбільш нестабільні умови довкілля і відповідно нестійкий гомеостаз у надорганізмових живих систем.
Від чого залежать темпи еволюції?
Темпи еволюції - проміжки часу, за який виникають певні систематичні групи (види, роди, родини і т. д. аж до типів тварин чи відділів рослин і царств). Під час створення синтетичної гіпотези еволюції темпи еволюційних процесів учені почали пов’язувати зі швидкістю зміни поколінь (мал. 39.7). Такі погляди базуються на таких міркуваннях: високі темпи зміни поколінь підвищують швидкість поширення мутацій у популяціях. Це призводить до змін фенотипів та врешті-решт - до утворення нових видів. Тобто швидкість зміни поколінь прямо пропорційна темпам еволюції.
Однак виявилося, що дані палеонтології не відповідають цій гіпотезі. Наприклад, у балтійському бурштині віком близько 40 млн років виявили 15 видів комах і кліщів, які практично не відрізняються від сучасних. Один з видів рачків-щитнів у викопному стані відомий починаючи з тріасового періоду мезозойської ери, тобто 230 млн років тому (мал. 39.8). Цим організмам властива висока швидкість зміни поколінь (не менш ніж раз на рік). З іншого боку, ряд хоботних, зміна поколінь у яких відбувається повільно (20-30 років), виник приблизно 36 млн років тому, дав у минулому велике число форм, а нині близький до вимирання. Отже, темпи еволюції та час існування певного виду не залежать від частоти зміни поколінь.

Мал. 39.7. Джордж Сімпсон (1902-1984) - американський палеонтолог та еволюціоніст. Встановив, що темпи еволюції різних систематичних груп залежать від швидкості зміни умов довкілля. Також досліджував форми добору

Мал. 39.8. Щитень літній - невеликий рачок (3-4 см завдовжки), мешканець прісних водойм, зокрема калюж. Без видимих змін деякі види щитнів існують починаючи з тріасового періоду мезозойської ери, тобто близько 230 млн років
Яке значення має біогеографія для розвитку еволюційного вчення?
Учені встановили, що різні частини біосфери розрізняються своїми групами ендемічних видів та біогеоценозами. Ендеміки (від грец. ендемос - місцевий) - види, роди, родини тощо, що мешкають лише в певній обмеженій частині біосфери (мал. 39.9). Ботанічне (флористичне) та зоологічне (зоогеографічне) районування не цілком збігаються внаслідок здатності тварин до активних міграцій, значного впливу властивостей ґрунту на рослини тощо.
Дані біогеографії використовують для обґрунтування положення про географічну ізоляцію як необхідну умову видоутворення. Однак нині зібрано свідоцтва про те, що ізоляція не завжди спричиняє дивергенцію та видоутворення. Так, більшість видів прісноводних і ґрунтових одноклітинних тварин має всесвітнє поширення. Одні й ті самі види ногохвісток (мал. 39.10) населяють усі континенти. Незважаючи на нездатність цих організмів до міграцій чи поширення іншими способами, їхні ізольовані одна від одної популяції не зазнають дивергенції. Отже, екологічна ніша виду зберігає його цілісність незалежно від можливості обміну спадковою інформацією між популяціями, а також форм розмноження.

Мал. 39.9. 1. Синьокрилий павич - ендемік тропічної Азії. 2. Баобаб, або адамсонія, - ендемічна рослина тропічної Африки. 3. Кролячий бандикут - невелика сумчаста тварина (до 50 см завдовжки, масою до 3 кг), мешканець пустель Австралії. 4. Велетенська черепаха - ендемік о-ва Альдабра біля західноафриканського узбережжя. 5. Плямиста, або вогняна, саламандра - ендемік Південної Європи; поширений у Львівській та Закарпатській областях України. 6. Морфоретенор - ендемік Південної Америки. Це великий метелик із переливчастим металево-блискучим забарвленням

Мал. 39.10. Ногохвістки - дрібні безкрилі членистоногі, здебільшого трапляються у ґрунті та підстилці
Дані біогеографії підтверджують єдність походження флори і фауни певних царств, областей, провінцій тощо. Наприклад, в областях Голарктичного царства від тундри до субтропіків існують подібні біогеоценози, до складу яких входять близькі види, наприклад звичайний та американський клени, європейська та американська норки (мал. 39.11) тощо. Це свідчить про існування суходільних зв’язків між Північною Америкою та північними континентами Східної півкулі в недалекому минулому. Дійсно, геологи довели, що протягом різних періодів та епох кайнозойської ери Північна Америка неодноразово з’єднувалась з Європою перешийком (його залишки - Гренландія та Ісландія) та з Азією - через суходіл на місці сучасної Берингової протоки.
Які причини вимирання видів?
Сучасний синтез даних біогеографії та геології дав змогу зробити висновки щодо причин вимирання певних груп організмів і бурхливої адаптивної радіації видів, які зайняли їхні місця. Багато вчених вважає, що так звані примітивні групи витісняються більш високоорганізованими «прогресивними». По суті, це подальший розвиток учення Ж.-Б. Ламарка про градації.

Мал. 39.11. Близькі види, які мешкають на території Євразії та Північної Америки: 1. Листок звичайного клена. 2. Листок американського клена. 3. Європейська норка. Невелика (від носа до хвоста до 65 см завдовжки) тварина родини кунячих. Мешкає в норах на берегах водойм Європи та Західної Азії. Занесена до Червоної книги України. 4. Американська норка зі своїм дитинчам. За способом життя нагадує європейську, однак трохи більша за розмірами (до 80 см завдовжки). Акліматизована в Європі, де витісняє норку європейську. Її розводять на звірофермах як хутрову тварину
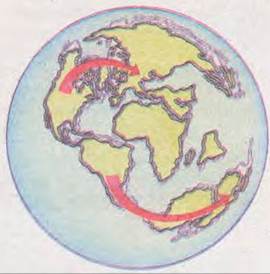
Мал. 39.12. Стрілками показано напрямок розселення сумчастих наприкінці крейдяного періоду (подано тогочасні обриси суходолу)
Наприклад, Ч. Дарвін вважав, що сумчасті ссавці були розповсюджені спочатку на всьому суходолі, однак згодом їх витіснили більш «прогресивні» плацентарні. В Австралії ж сумчасті збереглись завдяки її ізольованості від інших материків, яка зашкодила проникненню на її територію плацентарних. Дослідження американського вченого Л. Маршалла (1980) повністю спростували це твердження. Як свідчать дані палеонтології, сумчасті виникли в Америці в другій половині крейдяного періоду (близько 100 млн років тому). У кінці цього періоду Північна і Південна Америки втратили суходільний зв’язок, тому еволюція сумчастих у цих двох частинах світу відбувалась різними шляхами. На початку кайнозойської ери Північна Америка була з’єднана з Європою, куди і розселились сумчасті (мал. 39.12). Вони мешкали на цих континентах паралельно з плацентарними аж до антропогенового періоду, коли зникли внаслідок кількох послідовних зледенінь Північної півкулі.
Приблизно 12 млн років тому значне зледеніння відбулось у Південній півкулі, що призвело до зникнення австралійської біоти. Трохи раніше через суходільний місток, що існував між Південною Америкою та Австралією, через не вкриту ще тоді льодом Антарктиду до Австралії мігрували сумчасті. Вони пережили наступне зледеніння в північних місцевостях Австралії і згодом заселили весь материк, зазнавши широкої адаптивної радіації. Плацентарні (за винятком мишоподібних гризунів і рукокрилих) не змогли освоїти цю територію, бо суходільні адаптаційні зони ссавців вже були зайняті сумчастими.
Отже, основною причиною вимирання певних груп організмів є руйнування біогеоценозів, до складу яких вони входять. Насамперед вимирають високоспеціалізовані види з низькою екологічною (еволюційною) пластичністю.
При заселенні нових ділянок (наприклад, щойно виниклих островів) чи місць зруйнованих екосистем важливе значення має послідовність потрапляння до них певних груп організмів. Перші з них через відсутність конкуренції з екологічно близькими видами повністю освоюють певну адаптаційну зону, формуючи максимально можливу кількість екологічних ніш. Наприклад, дослідження фауни комах геологічно дуже молодих (виникли близько мільйона років тому) на Гавайських островах показали, що групи, які першими заселили ці території, дали величезну кількість ендемічних видів (один рід жуків-вусачів - 136 видів, мухи дрозофіли — понад 300 видів та ін.).
Нові терміни та поняття. Макроеволюція, біологічні прогрес і регрес, ароморфоз, загальна дегенерація, ідіоадаптація, темпи еволюції, ендеміки.
Запитання для повторення: 1. Що таке макроеволюція? 2. Як проявляються біологічні регрес і прогрес? 3. Що таке ароморфоз? Наведіть приклади. 4. Що таке загальна дегенерація? Наведіть приклади. 5. Що таке ідіоадаптація? Наведіть приклади. 6. У чому полягає інтеграція живого на різних рівнях його організації? 7. Від чого залежать темпи еволюції? 8. Яке значення біогеографії для доказів існування еволюції? 9. У чому причини вимирання видів?
Проблемне завдання. Поміркуйте, який характер (ароморфоз, ідіоадаптація, загальна дегенерація) мають такі еволюційні події: виникнення еукаріотів; зникнення крил у бліх та вошей; перетворення крил на ласти в пінгвінів; видозміна кореня на коренеплід; утворення квітки в покритонасінних.
Коментарі (0)