Гени
- 11-11-2021, 23:56
- 1 310
9 Клас , Біологія 9 клас Шаламов, Носов, Литовченко, Каліберда
§ 26. Гени
Дати визначення поняття «ген» не просто
У попередніх параграфах ми розглянули, як інформація про будову ДНК послідовно реалізується в процесах транскрипції, а потім трансляції. Спочатку РНК-полімераза синтезує матричну РНК на молекулі ДНК, а потім рибосома синтезує білок, зчитуючи інформацію з матричної РНК. В еукаріотів одна матрична РНК зазвичай містить інформацію про будову цілого типу білкових молекул. Ця матрична РНК синтезується з особливої ділянки ДНК, що має назву ген.
Отже, інформація про послідовність амінокислотних залишків у молекулі білка закодована в особливій ділянці молекули ДНК — гені. Проте гени кодують не тільки матричні РНК. Транспортні РНК, рибосомальні РНК та інші типи РНК також кодуються своїми генами, однак ці РНК ніколи не піддаються трансляції, залишаючись назавжди РНК. Таким чином, ми можемо сформулювати ширше визначення гена: ген — це ділянка ДНК, що кодує молекулу РНК1.
Розглянемо будову звичайного гена, що кодує матричну РНК тваринної клітини (рис. 26.1). Ген, як ми знаємо, є ділянкою дволанцюгової ДНК і лише один із ланцюгів кодує мРНК. Цей ланцюг називають антизмістовним. Послідовність нуклеотидів у ньому комплементарна матричній РНК, саме із цим ланцюгом працюватиме РНК-полімераза. Інший ланцюг ДНК за послідовністю нуклеотидів збігається з матричною РНК, що кодується геном. Цей ланцюг називають змістовним. При цьому один ланцюг ДНК може бути антизмістовним для одних генів і змістовним — для інших.

Тепер розглянемо, як розташовуються різні ділянки гена вздовж молекули ДНК. Ген починається з ділянки, що відповідає за зв’язування РНК-полімерази та початок транскрипції, — промотора. Слідом за промотором розташована ділянка гена, що кодує матричну РНК, — кодувальна частина гена. Як ви пам’ятаєте, у матричній РНК має бути старт-кодон. Такий само старт-кодон є й у гені. За ним розташована послідовність нуклеотидів, що кодує амінокислоти в білку.

Рис. 26.1. Будова гена еукаріотів
1 Проте навіть таке визначення недосконале. Під нього не підпадають, наприклад, гени вірусів із РНК-геномом. Також стало ясно, що часто в еукаріотів з одного гена синтезується кілька різних молекул РНК. У прокаріотів кілька генів можуть кодувати одну молекулу РНК, про що йтиметься в цьому параграфі. Хоча наше визначення поняття «ген» досить стисле, воно розкриває його суть.
Урешті-решт, кодуювальна ділянка закінчується стоп-кодоном1. Проте навіть після стоп-кодону транскрипція мусить якийсь час продовжуватися: молекула матричної РНК містить довгу ділянку, яка не транслюється рибосомою, але є важливою для функціонування матричної РНК. Ген закінчується ділянкою, що відповідає за припинення транскрипції, — термінатором. Термінатор забезпечує від’єднання РНК-полімерази від ДНК та припинення транскрипції. Як і у випадку промотора, термінатор може бути по-різному влаштований у різних генах. Цікаво, що послідовність, яка виконує роль термінатора в генах матричної РНК еукаріотів, не припиняє транскрипцію миттєво: РНК-полімераза зчитує ще 100-200 нуклеотидів і тільки тоді «відпадає» від ДНК. Зайві нуклеотиди потім відрізаються ферментами клітини.
Гени прокаріотів та еукаріотів відрізняються за будовою
Як ми вже зазначали, один із ланцюгів ДНК, а саме антизмістовний, є повністю комплементарним молекулі РНК, що синтезується на ньому РНК-полімеразою. Це означає, що якщо в пробірці ми додамо до цього ланцюга синтезовану на ньому РНК, то вони сформують дволанцюгову структуру на зразок тієї, яку формують два ланцюги ДНК. Але якщо ми додамо антизмістовний ланцюг ДНК до зрілої матричної РНК, що кодується цим геном, то виявиться, що кількість комплементарних ділянок буде не така вже й велика. Більша частина антизмістовної ДНК не буде комплементарна матричній РНК і формуватиме петлі. А комплементарна частина буде у вигляді кількох розрізнених ділянок (рис. 26.2).
Це пояснюється тим, що в еукаріотів після синтезу попередника матричної РНК із нього вирізаються невеличкі ділянки. Ці ділянки називають інтронами. Інтрони здебільшого не кодують амінокислотну послідовність білка. Вони безповоротно видаляються з молекули РНК у процесі її дозрівання. При цьому кодувальні ділянки, що залишилися, — екзони — з’єднуються між собою. Процес вирізання інтронів здійснюється в ядрі за участю малих ядерних РНК. Цей процес є невіддільною частиною дозрівання матричної РНК. Неймовірно, але інколи інтрони становлять до 90 % усієї послідовності гена! Виходить, що в ході дозрівання попередник мРНК укорочується в десять разів. Функції інтронів не до кінця вивчені. Їх уважають нащадками шкідливих послідовностей ДНК, що інфікували геном і безконтрольно розмножились у ньому (так звана егоїстична ДНК). За цією теорією вирізання інтронів — процес очищення генів від некодувальних послідовностей, що заразили їх. Проте клітини навчилися використовувати інтрони для того, аби кодувати кілька білків одним геном, збільшуючи цим різноманіття власних білків. Тому іноді інтрони все ж кодують ділянки амінокислотної послідовності білка. Припустімо, що в процесі вирізання один інтрон залишиться. Тоді інформація, що міститься в ньому, буде використана рибосомою для побудови молекули білка.

Рис. 26.2. Зв’язування зрілої мРНК та антизмістовного ланцюга ДНК
А. Схема. Б. Мікрофотографія.
1 В еукаріотів може бути багато стоп-кодонів. Зазвичай після головного розташовані кілька додаткових, щоб напевно припинити синтез білка. Більше того, як ми побачимо далі, у гені еукаріотів є ділянки, які будуть вирізані в процесі дозрівання матричної РНК. Ці ділянки не повинні транслюватися й часто містять стоп-кодони, аби запобігти синтезу неправильного білка в разі порушення їх вирізання.
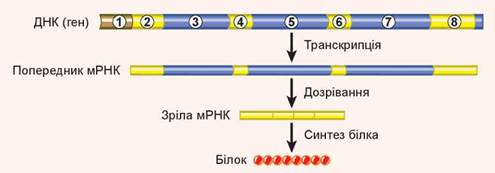
Рис. 26.3. Функціонування еукаріотичного гена
1. Промотор. 2, 4, 6, 8. Екзони. 3, 5, 7. Інтрони.
Цікаво, що в результаті цього процесу з одного гена утворюються різні матричні РНК, а з них — різні білки. Прикладом такого гена може бути ген фібронектину. Він містить фрагмент, що кодує ділянку РНК, яка в більшості клітин не вирізається, тобто розпізнається як екзон. Ця ділянка відповідає за зв’язування з білками міжклітинного матриксу, і її включення до молекули білка веде до формування нерозчинної форми фібронектину. Проте в клітинах печінки ця ділянка розпізнається як інтрон і вирізається. Це веде до формування циркулювальної форми фібронектину, розчиненої в плазмі крові. Таким чином, один ген кодує два білки. Вважається, що в людини так функціонує більшість генів, тому кількість білків у рази перевищує кількість генів, що кодують білки. Інтрони мають досить стале положення в генах, яке мало змінюється в процесі еволюції. Таким чином, гени, що кодують матричну РНК еукаріотів, мають уривчасту будову (рис. 26.3).
Ген прокаріотів значно відрізняється від еукаріотичного гена за будовою. По-перше, у прокаріотичних генах немає інтронів. При цьому вся послідовність гена транскрибується безпосередньо до матричної РНК. По-друге, одна молекула матричної РНК зазвичай кодує кілька білків. Ділянка ДНК, що кодує цю мРНК, має назву оперон (рис. 26.4). При цьому всі гени, із яких побудований оперон, мають лише один промотор на початку оперону. Оперон об’єднує кілька генів білків, залучених до спільного процесу й тому синтезованих разом. Найвідомішими є лактозний оперон, що кодує три білки, які беруть участь у споживанні лактози, і триптофановий оперон, що кодує п’ять білків, які беруть участь в біосинтезі амінокислоти триптофану. Більшість генів прокаріотів об’єднані в оперони. Цікавим є те, що зрідка оперони трапляються і в еукаріотів.
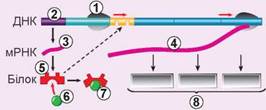
Рис. 26.4. Будова лактозного оперону
1. РНК-полімераза. 2. Ген білка-регулятора. 3. мРНК білка-регулятора. 4. мРНК трьох ферментів травлення лактози. 5. Білок-регулятор. 6. Лактоза. 7. Неактивний білок-регулятор. 8. Ферменти травлення лактози. Зверніть увагу, що білок-регулятор може заблокувати доступ РНК-полімерази до генів ферментів тільки за відсутності лактози в клітині.
Поміркуймо
Знайдіть одну правильну відповідь
1. Ген еукаріотів містить інформацію про
- А мРНК
- Б мРНК чи тРНК
- В тРНК чи рРНК
- Г мРНК, тРНК чи рРНК
- Д мРНК, тРНК, рРНК чи ДНК
2. Якщо матрична РНК має послідовність нуклеотидів АЦУГГЦА, то змістовний ланцюг ДНК мав послідовність
- А АЦУГГЦА
- Б УГАЦЦГУ
- В АЦГГТЦА
- Г АЦТГГЦА
- Д ТГАЦЦГТ
3. Правильне розташування елементів гена таке:
- А старт-кодон - промотор - кодувальна частина - стоп-кодон - термінатор
- Б промотор - старт-кодон - кодувальна частина - стоп-кодон - термінатор
- В старт-кодон - кодувальна частина - промотор - стоп-кодон - термінатор
- Г промотор - старт-кодон - кодувальна частина - термінатор - стоп-кодон
- Д старт-кодон - промотор - кодувальна частина - термінатор - стоп-кодон
4. В оперон об’єднуються гени, що кодують
- А білки мембран
- Б білки, потрібні для біосинтезу білка
- В різні РНК
- Г білки, потрібні для одного процесу
- Д рРНК
5. Еукаріоти вирізають інтрони під час дозрівання мРНК, тому що
- А рибосомі незручно працювати з довгою мРНК
- Б довгі мРНК не можуть залишити ядро крізь ядерну пору
- В інтрони несуть інформацію про шкідливі білки
- Г інтрони не кодують частини білка
- Д інтрони перешкоджають зчитуванню екзонів
Сформулюйте відповідь кількома реченнями
6. Чому для синтезу мРНК використовується антизмістовний ланцюжок ДНК? Чи можна зчитати той самий ген зі змістовного?
7. Довжина гена завжди більша за довжину недозрілої мРНК. Що є причиною цього? Куди зникають «зайві» фрагменти?
8. Чому при змішуванні дозрілої мРНК та антизмістовного ланцюга ДНК утворюються петлі?
9. Чого в клітині більше — генів чи різних видів білків, за ними синтезованих? Чи однаковою буде відповідь стосовно еукаріотів і прокаріотів та чому?
10. Чи важливим є те, у якому напрямку зчитувати ген в ході транскрипції? Як ви гадаєте, чим визначається напрямок зчитування?
11. Що спільного між Термінатором у виконанні Арнольда Шварценеггера в однойменному фільмі та термінатором гена?
Знайди відповідь і наблизься до розуміння природи
12. Оціни переваги та недоліки оперонної системи організації генів у прокаріотів. Чому еукаріоти «відмовилися» від такої системи?
13. Яку роль виконують малі ядерні РНК у процесі дозрівання мРНК?
Дізнайся самостійно та розкажи іншим
14. Крім генів, що кодують ферменти, до складу оперонів входять і регуляторні гени. Опиши роботу такої регуляції на прикладі лактозного оперону бактерій.
15. Альтернативний сплайсинг значно розширює різноманіття білків організму. У чому суть цього процесу та до яких захворювань може призводити його порушення?
Доповнення VII

Антон Абизов
Закінчив Харківський фізико-математичний ліцей № 27 у 2005 році. Переможець всеукраїнських олімпіад і турнірів з біології. Ступінь бакалавра з фізики отримав в Московському фізико-технічному інституті, ступінь магістра з біології в Еколь Політехнік (Франція). Дисертацію на здобуття наукового ступеня доктора філософії захистив в Університеті Гренобль-Альпи. Зараз працює в Центрі дослідження запальних процесів INSERM у Франції.
Гіпотеза «один ген — один білок»
Раніше вчені вважали, що кожному білку відповідає своя ділянка ДНК, яка має назву ген, і що один ген кодує один білок. Але вже наприкінці минулого століття виявилося, що один ген може кодувати кілька білків одразу, а для деяких ділянок ДНК кінцевим функціональним продуктом є навіть не білок, а молекула РНК.
Молекула РНК, отримана в ході транскрипції ДНК, не відразу бере участь у трансляції та синтезі поліпептидного ланцюга нового білка, а спершу зазнає істотної реорганізації — дозрівання. Щойно синтезована під час транскрипції молекула РНК — транскрипт — містить у собі екзони та інтрони. У ході дозрівання молекули РНК, окрім усього іншого, відбувається її сплайсинг: інтрони вирізаються, а екзони зшиваються разом, і така вже дозріла матрична РНК (мРНК) є основою для трансляції. За процес сплайсингу РНК у клітині відповідає цілий комплекс білків і молекул РНК — сплайсосома.

Рис. VII.1. Схема альтернативного сплайсингу гена з п’ятьма екзонами
У ході транскрипції синтезується ланцюг РНК, що містить інтрони та всі п’ять екзонів, але до зрілої мРНК потрапляють лише деякі екзони, тож у результаті виникає три варіанти мРНК, що відповідають трьом формам білкового продукту.
Але не завжди всі екзони зшиваються та потрапляють до мРНК. Деякі з них за певних умов у ході сплайсингу вирізаються разом з інтронами, а іноді деякі інтрони не вирізаються і залишаються в мРНК. У результаті цього явища, що має назву альтернативний сплайсинг, відбувається трансляція різних варіантів білка з того самого гена. Схематично процес альтернативного сплайсингу зображено на рисунку VII.1.
Альтернативний сплайсинг було відкрито наприкінці 1970-х рр. в аденовірусів, але його поширеність і значення для живих організмів вдалося зрозуміти лише нещодавно.
Зокрема, виходячи з гіпотези, що один ген кодує один білок, учені не могли пояснити, чому в людини лише 23 тисячі генів, що не набагато більше, ніж генів у геномі круглого черва ценорабдітіса (близько 20 000), але при цьому організм людини організований набагато складніше, ніж у круглих червів. Аналіз матричних РНК, проведений 2008 року вченими із США, показав, що альтернативному сплайсингу піддаються до 94 % генів людини. Водночас у ценорабдітіса альтернативний сплайсинг зачіпає лише близько 15 % генів. Виходить, що за подібної кількості генів в організмі людини синтезується набагато більше різних білків.
Найкраще альтернативний сплайсинг був вивчений у дрозофіли, яка впродовж десятиліть є улюбленим об’єктом різноманітних генетичних досліджень. Відмінності між самцями та самицями в цих плодових мушок визначаються каскадом взаємопов’язаних генів, що піддаються альтернативному сплайсингу. Наведемо приклад: стать у дрозофіл визначається кількістю X-хромосом, яка впливає на альтернативний сплайсинг гена Sxl. При цьому функціональний білок Sxl зчитується лише в самиць. Цей білок регулює альтернативний сплайсинг гена Tra, у результаті його дії відбувається синтез функціонального білка Tra, знову-таки, лише в самиць. Своєю чергою, у нервовій системі дрозофіл білок Tra регулює альтернативний сплайсинг гена Fru, але вже таким чином, що білок на отриманій мРНК у самиць не утворюється. У підсумку функціональний білок Fru є лише в самців, і саме він визначає поведінкові навички, що потрібні їм для залицяння до самиць! У ході експерименту було доведено, що самиці-мутантки із «чоловічим» варіантом білка Fru теж залицяються до самиць.
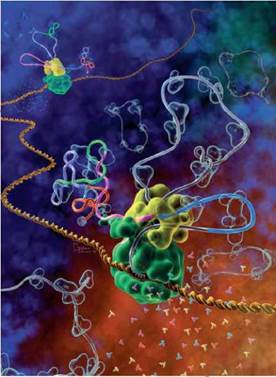
Рис. VII.2. Альтернативний сплайсинг очима художника
Дві РНК-полімерази (зелений глобулярний білок) рухаються подвійною спіраллю ДНК (жовтогарячого кольору) у глибину рисунка, синтезуючи молекулу мРНК — транскрипт (зображений як стрічка). Синтезований транскрипт РНК містить різні екзони (кольорові ділянки) та інтрони (сірі ділянки стрічки). Сплайсосома (жовта глобулярна структура) вирізає черговий інтрон (сіра петля вгору) і з’єднує кінці синього та рожевого екзонів. У вже опрацьований ланцюг РНК (ліворуч від сплайсосоми) включений зелений екзон. А друга сплайсосома (у глибині рисунка), навпаки, вирізає зелений екзон зі свого ланцюга РНК. Вирізані інтрони, замкнені в кільця, пливуть геть від сплайсосоми.
Коментарі (0)