Популяційна генетика людини
- 30-07-2021, 22:18
- 1 114
10 Клас , Біологія і екологія 10 клас Остапченко (рівень стандарту)
§ 38. ПОПУЛЯЦІЙНА ГЕНЕТИКА ЛЮДИНИ
Пригадайте, що таке популяція, вид, ареал виду, геном, мутації. Що собою становлять мікросателітні повтори?
Генетика популяцій. Людина є одним з найпоширеніших і найчисленніших видів на Землі. Можна сказати, що ареалом нашого виду є майже вся поверхня планети. Як будь-який вид, усю сукупність особин нашого виду можна поділити на популяції — локальні, почасти відокремлені групи осіб. Популяції, як і окремі організми, мають чіткі індивідуальні відмінності і є динамічними системами. Кожну популяцію можна описати за різними параметрами: ареал, чисельність, густота, вікова та статева структура, народжуваність, смертність, приріст, екологічними та генетичними характеристиками. Ми приділимо увагу параметрам, які характеризують генетичну структуру популяції, її генофонд.
Генетичну структуру популяції (та процеси, що призводять до її зміни) вивчає окремий розділ генетики — генетика популяцій. Сукупність усіх генів та їхніх алелів у популяції називають генофондом. Про особин, які належать до однієї популяції, кажуть, що вони мають спільний генофонд, оскільки в результаті вільних схрещувань у популяції відбувається і вільний обмін генетичною інформацією.
Дослідження процесів успадкування і мінливості в межах популяції мають свої особливості. Насамперед — це потреба враховувати всі генетичні наслідки від всієї кількості вільних схрещувань, що відбуваються у популяції. Крім того, популяції більш генетично різноманітні, ніж окремі індивідууми (чи родина). У геномі людини як диплоїдного організму будь-який ген може бути представлений максимум двома різними алелями. Але в людських популяціях таких алелів може бути багато, тому можна зустріти різні варіанти генотипів і, відповідно, різні варіанти фенотипу. Яскравий приклад — групи крові за системою АВ0 (пригадайте явище множинного алелізму та як успадковуються групи крові за системою АВ0 в людини). Що більше різних алелів певних генів представлено в популяції, то різноманітнішим буде її генофонд. У разі змін умов існування це дає змогу «мобілізувати» весь резерв спадкової мінливості (комбінативної та мутаційної) для формування відповідних адаптацій. Отже, різноманітний генофонд популяції підвищує її адаптивний потенціал. Пригадайте: адаптивний потенціал — міра пристосувальних можливостей популяції або виду до умов середовища.
Визначення гетерогенності популяції. Якщо в популяції спостерігаємо тільки один стан ознаки, то така ознака є мономорфною. Наявність у популяції різних станів ознаки (а відповідно, і певного гена) називають поліморфізмом (мал. 38.1). Отже, поліморфність популяції можна визначити як частку поліморфних генів (чи ознак, які вони зумовлюють) серед усіх проаналізованих особин.

Мал. 38.1. Поліморфізм за кольором очей у людини
У популяційній генетиці поліморфним вважають алель, частота зустрічальності якого в популяції становить 1 % та більше. Алелі, частота зустрічальності яких не досягає критерію поліморфізму, але значно вища за рівень спонтанних мутацій, називають рідкісними. Явище поліморфізму свідчить про те, що в особин цієї популяції частина генів перебуває у гетерозиготному стані.
Що більша кількість алелів відповідного гена в популяції, то більше в ній осіб, які є гетерозиготами. Частота гетерозигот за відповідним локусом у даній популяції є другим показником її генетичної різноманітності — гетерозиготності. Головними параметрами генетичної структури популяції (її генофонду) є частоти зустрічальності алелів (концентрація їх у популяції) та певних генотипів.
Визначення частоти зустрічальності алелів і генотипів у популяції. Частоту зустрічальності відповідного алеля визначають як відношення кількості цього алеля в популяції (враховуючи, що кожна диплоїдна гомозигота має по два подібні алелі певного гена, а гетерозиготи — по одному) до загальної кількості алелів цього гена в популяції. У разі повного домінування (гетерозиготи фенотипово не відрізняються від домінантних гомозигот) прямий розрахунок частоти зустрічальності алелів і певних генотипів неможливий. Її можна визначити, виходячи із закону Гарді — Вайнберга, який у 1908 р., незалежно один від одного, сформулювали Ґ. Гарді та В. Вайнберг (мал. 38.2).

Мал. 38.2. Ґодфрі Гарольд Гарді (1877—1947) — британський математик (А) та Вільгельм Вайнберг (1862—1937) — німецький лікар (Б) незалежно один від одного сформулювали закономірність розподілу алелів і генотипів у популяції
Запам'ятаємо
Закон Гарді — Вайнберга полягає в тому, що в достатньо численній популяції, у якій відбувається вільне схрещування особин, відсутній вплив будь-якого зовнішнього фактору, який сприяє чи не сприяє певним поєднанням алелів, не виникають нові мутації, не відбувається обмін генетичною інформацією з іншими популяціями (наприклад, унаслідок міграцій особин), співвідношення алелів стабілізується протягом кількох поколінь і тривалий час залишається сталим.
Співвідношення генотипів нащадків гетерозиготних батьків у численній популяції, де відбувається вільне схрещування, наведено в таблиці 38.1 (p — частота домінантного алеля, q — частота рецесивного алеля).
Таблиця 38.1
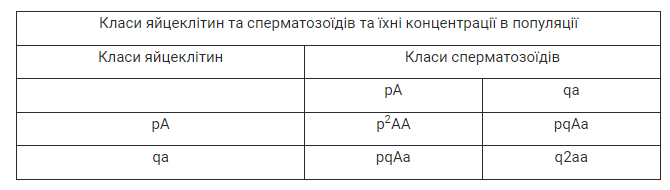
Із цієї таблиці випливає, що розподіл генотипів нащадків визначається за формулою:
р2АА + 2pqAa + q2aa = 1 (тобто 100 %),
або у скороченому вигляді:
(pA + qa)2 = 1.
Оскільки ми припустили, що є лише два алелі певного гена, то зрозуміло, що:
pA + qa = 1.
Сума частот усіх алелів, як і сума частот усіх генотипів, дорівнюватиме 1. Однак наведена формула як раз і описує ситуацію, коли дійсно різні генотипні класи нащадків траплятимуться з такою самою частотою, як і в батьків (рівноважне співвідношення). Явище підтримання такого співвідношення називають генетичною рівновагою у популяції. Рівновага зберігається і тоді, коли певний ген представлений не двома, а більшою кількістю алелів.
Цікаво знати
Формула, яка описує закон Гарді — Вайнберга, є не що інше, як перенесений на групу особин, де обов'язково є вільне схрещування, результат, який описує другий закон Менделя.
Як застосовують закон Гарді — Вайнберга? Коли досліджують розподіл частот генотипів у популяції при успадкуванні аутосомно-рецесивної або аутосомно-домінантної ознаки, то за фенотипом можна точно з’ясувати кількість гомозигот за рецесивним алелем: їхній генотип визначатиме лише один варіант фенотипу. Але відрізнити гомозигот за домінантним алелем від гетерозигот за фенотипом неможливо. У такій ситуації застосувують закон Гарді — Вайнберга.
Розглянемо приклад. В Україні приблизно серед 5000 людей можна знайти одну людину з фенілкетонурією (неможливість метаболізувати амінокислоту фенілаланін, що за відсутності спеціальних заходів призводить до важких порушень нервової системи). Хвороба успадкується як аутосомно-рецесивна. Потрібно визначити відсоток осіб у популяції, які є носіями патологічного рецесивного алеля (поміркуйте, навіщо взагалі визначати частоту носіїв патологічних генів у популяції). Основна проблема полягає в тому, що серед здорових осіб носіїв патологічного рецесивного алеля (вони є гетерозиготами) за фенотипом виявити неможливо. Зробити це можна, використовуючи формулу, яку описує закон Гарді — Вайнберга. Ми знаємо, що частота гетерозигот — це подвоєний добуток частот домінантного та рецесивного алелів, тобто спочатку нам потрібно знайти частоти рецесивного та домінантного алелів. Легко знаходимо частоту гомозигот за рецесивним алелем: q2 = 1/5000 = 0,0002
Відповідно частота рецесивного алеля становитиме: q = √0,0002 = 0,014
Частота домінантного алеля становитиме: P = 1 - q = 1 - 0,014 = 0,986.
Частота гетерозигот дорівнюватиме: 2pq = 2 х 0,014 х 0,986 = 0,276, або 2,76 %.
Класичний математичний запис закону Гарді — Вайнберга (але не сам закон!) справедливий виключно для аутосомних генів. Для генів, що містяться у статевих хромосомах, логіка визначення частот зустрічальності алелів і генотипів буде дещо іншою.
Розглянемо приклад. Часто виникає питання, чому за Х-зчепленого рецесивного успадкування в родоводах немає жінок з хворобами. Відомо, що найпоширеніша форма гемофілії (успадковується як Х-зчеплена рецесивна ознака) трапляється серед осіб чоловічої статі із частотою 1 людина з хворобою на 2500 чоловіків. Скільки ми можемо очікувати осіб із хворобою і носіїв патологічного гена серед жінок? Пригадаємо, що чоловіки за генами, які містяться в Х-хромосомі, є гемізиготами (тобто в Y-хромосомі відповідні алелі відсутні), а жінки, на відміну від чоловіків, можуть бути як гомозиготами, так і гетерозиготами. Частоти домінантного та рецесивного алелів відповідного гена ми можемо легко з'ясувати, виходячи з кількості чоловіків із хворобою і здорових: популяційна частота алеля Х-зчепленого гена дорівнюватиме частоті чоловіків, у яких спостерігають фенотиповий прояв відповідного алеля (обґрунтуйте чому). Таким чином, частота рецесивного алеля, що спричинює гемофілію: qh = 1/2500 = 0,0004, а домінантного: pH = 1 - 0,0004 = 0,9996. Відповідно, серед жінок частота з хворобою (гомозигот за рецесивним алелем) дорівнюватиме (qh)2 = 0,00042 = 0,00000016 (1 жінка з хворобою трапляється серед 6 250 000 жінок; навіть за масштабного генеалогічного дослідження ми не будемо мати такої кількості людей!). Частота жінок — носіїв мутантного алеля буде значно більшою: 2pHqh = 2 х 0,9996 х 0,0004 = 0,0008 (1 жінка-носій серед 1250 жінок).
Закон Гарді — Вайнберга справедливий лише для великих популяцій, де відбувається вільне схрещування та відсутні фактори (міграції, ізоляції, природний добір, мутаційна мінливість, популяційні хвилі), які змінюють частоти алелів і генотипів. Зрозуміло, що популяцій, які б відповідали всім переліченим вимогам, у природі не існує.
Людські спільноти чимось подібні до природних популяцій організмів, але дуже від них відрізняються. Так, у зв’язку з розвитком засобів пересування дедалі більше втрачається значення географічних перешкод в ізоляції певних груп людей. Масові міграції сприяють обміну генетичним матеріалом між спільнотами, що мешкають на значній відстані одна від одної. Унаслідок цього можуть поступово зменшуватись відмінності в генофонді людських спільнот, ізольованих географічно.
Якщо дві популяції істотно розрізняються за своєю генетичною структурою, це може істотно вплинути на зміну частоти алелів при змішуванні осіб цих популяцій. Неспоріднені шлюби між мігрантами та місцевим населенням супроводжуються підвищенням гетерозиготності. Тому в нащадків від таких шлюбів рецесивні алелі, що здатні в гомозиготному стані спричиняти певні спадкові захворювання, часто перебувають у гетерозиготному стані і тому не проявляються у фенотипі.
Порівнюючи розповсюдження генів і генотипів у різних популяціях, можна прослідкувати міграцію окремих людських угруповань, шляхи переселення цілих народів, а також зробити висновки про генетичну спорідненість народів (національностей), які проживають на суміжних територіях.
В ізоляції людських спільнот значна роль може належати соціальним факторам (мова, релігійні погляди, належність до певного народу чи нації, історичні традиції, соціальний стан, освіта тощо). Часто шлюби беруть особи з однаковими фізіологічними порушеннями (наприклад, слуху і здатності до мови), бо вони краще досягають взаєморозуміння та мають спільні інтереси. Унаслідок дії соціальних факторів люди, що проживають на спільній території, можуть утворювати обмежені групи — ізоляти.
Трапляються випадки, коли шлюб укладають близькі родичі (явище інбридингу) або люди, які належать до генетично віддалених ліній (наприклад, до різних національностей або навіть різних рас) — явище аутбридингу. Наслідком споріднених шлюбів є зростання серед нащадків частоти гомозигот за певним алелем. Оскільки багато летальних або сублетальних алелів рецесивні, у результаті інбридингу збільшується кількість рецесивних гомозигот і спостерігають явище інбредної депресїі — зниження життєздатності осіб. Це пояснюють тим, що при переході в гомозиготний стан летальних і сублетальних алелів патологічні ознаки (різноманітні захворювання або вроджені вади розвитку), які вони визначають, проявляються у фенотипі.
Сукупність шкідливих точкових (генних) і хромосомних мутацій у генофонді називають генетичним тягарем популяції.
Завдання. За допомогою різноманітних джерел інформації знайдіть приклади родоводів із близькоспорідненими шлюбами (наприклад, родовід іспанського короля Карла ІІ) та проналізуйте їх.
Кожна людина є носієм приблизно 300 алельних генів відомих спадкових хвороб. Небезпека народження дитини з певними спадковими захворюваннями або генетичними вадами зростає зі збільшенням ступеня спорідненості осіб, які беруть шлюб. Ступінь спорідненості характеризується коефіцієнтом інбридингу — імовірністю того, що в генотипі якоїсь особини будуть наявні два алелі, ідентичні за походженням, тобто походять від спільного предка. Так, для нащадків рідних братів і сестер цей коефіцієнт дорівнює 1/4 (це означає, якщо один з батьків має певний алель, то ймовірність того, що такий саме алель є і в іншого, становить 1/2), для двоюрідних братів і сестер — 1/16, троюрідних — 1/64.
На генофонд популяції у наступних поколіннях може впливати природний добір. Якщо певний варіант фенотипу, який визначається відповідним варіантом генотипу, виявиться несприятливим у даних умовах існування, то в таких особин, відповідно, менше шансів вижити й залишити плодючих нащадків. Вочевидь, що при вилученні з процесу розмноження певного генотипу, наприклад рецесивної гомозиготи, частота зустрічальності цього генотипу та, відповідно, рецесивного алеля буде зменшуватися впродовж наступних декількох поколінь. Таким чином, тиск добору може змінити співвідношення частот зустрічальності алелів у наступних поколіннях. Такий процес завжди відбувається у певному напрямку: надаючи перевагу одному варіанту генотипу, добір може повністю вилучити з популяції інший.
Хоча до природного добору залучені фенотипи особин, одиницею добору є генотип, тобто відповідний набір генетичної інформації. Тому що різноманітніший генофонд популяції, то ефективніша дія природного добору.
Природний добір і мутаційний процес «працюють» спільно. Якщо новоутворений варіант послідовності нуклеотидів у молекулах ДНК забезпечує розвиток адаптивної в даних умовах ознаки, такий алель «підхоплюється» добором і фіксується в наступних поколіннях.
Як ми вже згадували, для того щоб виконувався закон Гарді — Вайнберга, популяція має бути численною. Це забезпечує утворення всіх можливих гамет та їхніх комбінацій під час формування наступного покоління. Коли популяція нечисленна, то підвищується імовірність випадкового відхилення від середньої частоти зустрічальності якогось алеля, а це може призвести до істотної зміни частот алелів і генотипів.
Процес випадкової і неспрямованої зміни частот зустрічальності алелів у нечисленній популяції має назву дрейф генів. Найчастіше дрейф генів спостерігають за різкого зменшення чисельності популяції. Наприклад, коли внаслідок міграції невелика кількість особин переселяється на нову територію. У такому разі кажуть, що популяція наче проходить крізь «шийку пляшки»: у маленькій новій популяції зазвичай будуть представлені не всі алелі, притаманні вихідній популяції, а лише частина. Крім того, частота зустрічальності алелів у новоствореній популяції буде іншою порівняно з вихідною. Отже, генофонд наступних поколінь визначатиме невелика кількість особин новоствореної невеликої популяції (ефект засновника) (мал. 38.3). Одним з наслідків дрейфу генів є зростання частки гомозиготних особин (поміркуйте чому).
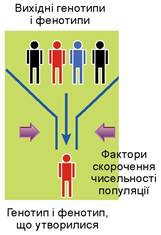
Мал. 38.3. Дрейф генів при швидкому скороченні чисельності популяції (ефект «шийки пляшки»)
Ключові терміни та поняття
генетика популяцій, адаптивний потенціал, закон Гарді — Вайнберга, генетичний тягар популяції, дрейф генів.
Перевірте здобуті знання
1. Які завдання стоять перед генетикою популяцій? 2. Що таке генофонд популяції? Чим він визначається? 3. Що таке адаптивний потенціал? Яка його генетична основа? 4. Сформулюйте закон Гарді — Вайнберга. За яких умов він виконується? 5. Як застосовують закон Гарді — Вайнберга в популяційній генетиці людини? 6. Що спільного та відмінного між природними популяціями та людськими спільнотами? 7. Що таке дрейф генів? Які його генетичні наслідки?
Поміркуйте
Чому дрейф генів меншою мірою впливає на генофонд численної популяції порівняно з нечисленною?
Коментарі (0)