Основні положення синтетичної гіпотези еволюції
- 16-05-2021, 13:30
- 817
11 Клас , Біологія і екологія 11 клас Балан, Вервес (рівень стандарту, академічний рівень)
§ 37. Основні положення синтетичної гіпотези еволюції
Аби краще засвоїти матеріал цього параграфа, слід пригадати: які закономірності спадкової мінливості відкрив Г. Мендель? Що зумовило кризу дарвінізму на початку XX століття? Що таке норма реакції? Що таке генофонд, фенотип і фени? Що таке підвид?
Невирішені проблеми загальної біології початку XX сторіччя дали поштовх для створення низки гіпотез, серед яких для подальшого розвитку еволюційних поглядів першорядне значення мали синтетична гіпотеза еволюції та гіпотеза неокатастрофізму.
У чому суть синтетичної гіпотези еволюції?
Синтетична гіпотеза еволюції - це комплекс понять про еволюційний процес, що склалися в 20-50-х роках XX сторіччя. Серед її творців були видатні вчені з різних країн світу (мал. 37.1). Її було розроблено як наслідок поєднання уявлень про мутації як єдине джерело спадкової мінливості, популяцію як елементарну одиницю еволюції та дарвінівських уявлень про боротьбу за існування та природний добір. Основні положення цієї гіпотези такі:
1. Єдиним джерелом спадкової мінливості є мутації.
2. Усі еволюційні перетворення відбуваються в популяціях - елементарних одиницях еволюції.

Мал. 37.1. Видатні вчені, які зробили вагомий внесок у розвиток синтетичної гіпотези еволюції: 1 - Сергій Сергійович Четвериков (1880-1959). Російський учений, засновник генетики популяцій. Розробив учення про популяцію як елементарну одиницю еволюції. Ввів термін «хвилі життя»; 2 - Феодосії Григорович Добржанський (1900-1981). Американський генетик. Закінчив Київський національний університет імені Тараса Шевченка і певний час працював у ньому. Зробив кілька відкриттів у галузі вивчення мутацій і мутагенезу. Досліджував основні особливості мікроеволюційних процесів. Засновник фенетики (від грец. файно - являю, знаходжу) - науки про прояв і розподіл фенів, тобто різних станів певної ознаки в популяції; 3 - Олексій Миколайович Сєверцов (1866-1936) - російський зоолог; з 1902 по 1911 р. працював професором у Київському університеті Св. Володимира (нині - імені Тараса Шевченка). Автор концепції про біологічний прогрес і шляхи його досягнення (ароморфози, ідіоадаптації, дегенерації). Розробив основи сучасної порівняльної анатомії хребетних тварин; 4 - Іван Іванович Шмальгаузен (1884-1963). Видатний український учений, спеціаліст у галузі еволюційної морфології та порівняльної анатомії та ембріології хордових тварин. Закінчив у 1907 р. Київський університет Св. Володимира (нині - імені Тараса Шевченка), згодом працював у ньому професором. У 1930-1941 рр. очолював Інститут зоології НАН України, який носить його ім’я
3. Елементарними факторами еволюції є боротьба за існування, хвилі життя, ізоляція, дрейф генів.
4. Існують три види еволюційного процесу: мікроеволюція, видоутворення та макроеволюція.
5. Рушійною силою еволюції є природний добір, який діє на сукупність фенотипів популяції. Природний добір буває стабілізуючим, рушійним і розриваючим (дизруптивним).
6. Будь-яка систематична група організмів може або процвітати (перебувати у стані біологічного прогресу), або вимирати (перебувати у стані біологічного регресу). Біологічний прогрес досягається за рахунок змін у будові: ароморфозів, ідіоадаптацій чи загальної дегенерації.
7. Процес еволюції необоротний, тобто при поверненні умов довкілля до попереднього стану адаптації кожного разу розвиваються заново, а не відтворюються попередні.
Чому популяцію вважають елементарною одиницею еволюції?
Синтетична гіпотеза еволюції розглядає популяцію як елементарну одиницю еволюції, бо в ній відбуваються всі основні еволюційні процеси: спадкова мінливість, боротьба за існування, природний добір, хвилі життя (популяційні хвилі), ізоляція, дрейф генів. Окремі особини, які складають популяцію, не мають власної еволюційної долі, бо всі еволюційні події здійснюються під час зміни поколінь.
Елементарними факторами еволюції, на думку творців синтетичної гіпотези, є хвилі життя, дрейф генів та ізоляція.
Пригадайте, хвилі життя (популяційні хвилі) - це коливання чисельності особин популяції, характерні для всіх видів. Вони можуть залежати від циклу розвитку виду і бути зумовлені генетично, наприклад сезонні коливання чисельності однорічних рослин, комах тощо, або викликані змінами інтенсивності екологічних факторів (умови зимівлі, прес хижаків, паразитів, наявність їжі тощо), мати періодичний чи неперіодичний характер (мал. 37.2).
Еволюційне значення хвиль життя полягає в тому, що при зменшенні або збільшенні чисельності особин частка тих чи інших алелів у генофонді популяції може змінюватись незалежно від ступеня пристосованості організмів. Наприклад, під час суворої зими вимерзають організми верхніх шарів ґрунту і залишаються лише ті з них, що випадково потрапили в місця, де замерзання менш інтенсивне (наприклад, у затишку, де вітер не здуває сніг, тощо). Таким чином, хвилі життя є одним з чинників, здатних спричиняти зміни генофонду популяцій.

Мал. 37.2. Хвилі життя можна простежити на прикладі перелітної сарани. У разі масового розмноження сарана збирається у велетенські зграї і перелітає з місця на місце, виїдаючи всю зелену рослинність на своєму шляху. Стимулом до гуртування слугує висока густота популяції. Якщо густота популяції невелика, то сарана веде поодинокий спосіб життя
Дрейф генів — випадкова та неспрямована зміна частот зустрічальностей певних алелів та їхніх поєднань у генофондах популяцій, яка не має пристосувального характеру. Найчіткіше це явище виявляється у нечисленних популяціях завдяки обмеженню вільного схрещування. І навпаки, чим вища чисельність популяції, тим менш значна роль дрейфу генів у зміні частот зустрічальності алелей та їхніх поєднань.
Розробив учення про дрейф генів як елементарний фактор еволюції американський генетик Сьюел Райт (1889-1982). Він виконав такі досліди. У пробірки з поживним середовищем С. Райт помістив по два самці та дві самки дрозофіли, гетерозиготні за певним геном (запишемо їхній генотип як Аа, при цьому нормальний алель - дикий тип - позначимо як А, а мутантний - як а). Отже, у цих штучних мікропопуляціях частоти зустрічальностей обох алелів дорівнювали 50 %. Через деякий час, за який змінилося декілька поколінь дрозофіл, виявили, що в деяких мікропопуляціях усі особини стали гомозиготними за рецесивним алелем а, в інших - за домінантним алелем А, а в інших збереглися обидва алелі, але співвідношення частот їхніх зустрічальностей відрізнялося від подібних показників на початку експерименту. Цей дослід показав, що в нечисленних популяціях частоти зустрічальностей алелів та їхніх поєднань можуть різко і неспрямовано змінюватись. При цьому може зберігатись мутантний алель, незважаючи на те, що він знижує пристосованість особин.
Однією з причин дрейфу генів є популяційні хвилі. Найчіткіше це виражене в організмів із швидкою зміною поколінь (наприклад, комах, кліщів, однолітніх рослин). При цьому густота популяції зменшується через загибель частини особин з випадкових причин, незалежно від ступеня їхньої пристосованості до умов довкілля, що значно змінює частоти зустрічальностей різних алелів та їхніх поєднань.
Отже, дрейф генів може мати такі наслідки. По-перше, в популяціях, особливо нечисленних, може зростати частота зустрічальностей гомозигот. Це зумовлено підвищенням ймовірності спорідненого схрещування, наслідком якого є зростання частки гомозигот. По-друге, у популяції тривалий час може зберігатись мутантний алель, який знижує пристосованість особин. По-третє, завдяки популяційним хвилям може швидко збільшуватись частота зустрічальностей одних алелів, тоді як інші взагалі можуть бути втрачені. Отже, унаслідок дрейфу генів популяції, які спочатку мали подібну генетичну структуру, можуть з часом усе більше за нею розрізнятись, незалежно від змін умов існування. Про інтенсивність дрейфу генів судять, виходячи із зростання частки гомозигот.
Дрейф генів як фактор еволюції має особливе значення при заселенні нових територій, наприклад островів. Зазвичай на них потрапляє невелике число особин одного виду з різними генотипами, і тому генофонд такої популяції з покоління до покоління все більше відрізнятиметься від генофондів вихідних популяцій. Під час дрейфу генів певні поєднання алелів виявляються корисними та зберігаються у нащадків; тому зміни генофондів мають пристосувальний характер.
Ізоляція - умови, за яких схрещування між особинами одного виду стає неможливим. Розрізняють кілька форм ізоляції. За географічної ізоляції різні популяції роз’єднані непрохідним для них географічним бар’єром. Наприклад, якщо річка чи гірський хребет виникають на території між двома популяціями рівнинного виду, нездатного плавати чи літати, коли один лісовий масив поділяється на два степом тощо.
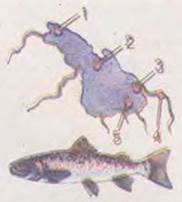
Мал. 37.3. Приклади екологічної ізоляції: 1-5 - місця нересту різних стад севанської форелі
Екологічна ізоляція відбувається, якщо всередині популяції виникають певні форми (раси), кожна з яких має відмінні вимоги до умов середовища життя. Часто виникають внутрішньовидові групи тварин, які відрізняються характером їжі. Наприклад, одні раси жука горохової зернівки живляться насінням гороху, інші - квасолі тощо. В період розмноження ці раси між собою не контактують завдяки різним місцезростанням кормових рослин.
Ізоляція може виникнути через різні строки розмноження особин одного виду в різних умовах. Наприклад, різні рослини лучних трав певного виду цвітуть залежно від часу весняної повені: особини, що довше перебували під водою, цвітуть пізніше від тих, які взагалі не були затоплені чи перебували під водою короткий час.
Ізоляція часто залежить від особливостей поведінки. Наприклад, в озері Севан (Вірменія) мешкає кілька стад форелі, які живляться разом, однак кожне стадо на нерест прямує в особливе місце (мал. 37.3).
Певні форми ізоляції можуть діяти незалежно одна від одної і водночас у різних популяціях одного виду. Еволюційне значення ізоляції полягає в тому, що за відсутності обміну алелями між популяціями певного виду в генофонді кожної з них виникають різні мутації та змінюються частоти зустрічальностей алелів, за рахунок чого популяції пристосовуються до умов довкілля незалежно одна від одної. Отже, ізоляція - це необхідна умова розходження ознак всередині одного виду.
Які відомі форми дії природного добору?
Залежно від спрямування адаптаційних змін організмів природний добір буває стабілізуючим, рушійним і розриваючим (дизруптивним) (мал. 37.4).
Стабілізуючий добір проявляє себе в постійних умовах довкілля. Він підтримує сталість певного фенотипу, найбільш відповідного навколишньому середовищу. Натомість відкидаються будь-які зміни, як менш адаптаційні, тим самим звужуючи норму реакції (тобто межі модифікаційної мінливості). Наприклад, два сучасні види кистеперих риб - латимерії (мал. 37.5) - дожили до наших днів, оскільки мешкають на значних глибинах, де умови існування досить постійні.
Рушійний, або спрямований, добір відбувається в разі змін у навколишньому середовищі або під час пристосувань до нових умов при розширенні ареалу. Він сприяє змінам мінливості в певному напрямі, зсуваючи у відповідний бік норму реакції. Наприклад, при пристосуванні тварин до риючого способу життя в різних неспоріднених групах виникають копальні кінцівки (вовчок, жук-гнойовик, сумчасті та плацентарні кроти тощо).
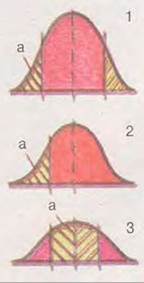
Мал. 37.4. Форми природного добору: 1 – стабілізуючий; 2 - рушійний; 3 - розриваючий (а - загибель особин)

Мал. 37.5. Два сучасні види латимерій

Мал. 37.6. Дія розриваючого (дизруптивного) добору: різні форми забарвлення сонечка двокрапкового; форми з червоними надкрилами (1) краще пристосовані до дії підвищених температур, а з чорними (2) - до дії низьких
Розриваючий, або дизруптивний, добір (від лат. дизруптус - розірваний) спрямовує мінливість у двох, рідше кількох, різних напрямах, однак не сприяє середнім (проміжним) станам ознак (мал. 37.6). Наприклад, у популяціях комах океанічних островів, де постійно дме сильний вітер, унаслідок дизруптивного добору є особини без крил або з дуже добре розвинутими крилами, які здатні протистояти здуванню, а комахи із середнім ступенем розвитку крил зникли. Отже, цей добір спричиняє виникнення кількох фенотипних форм у популяції (поліморфізму), що сприяє її пристосуванню до нестабільних умов довкілля.
Що таке мікроеволюція? Яке її значення для еволюції організмів?
Мікроеволюція - сукупність еволюційних процесів, які відбуваються в популяціях одного виду.
Згідно із синтетичною гіпотезою еволюції, єдиним джерелом спадкової мінливості є мутації. Природний добір спрямовує різні елементарні спадкові зміни фенотипів (фени), що виникли внаслідок мутацій, на шлях утворення адаптацій - пристосувань організмів до умов навколишнього середовища (мал. 37.7). У цьому полягає творча роль природного добору. Саме тому його часто називають рушійною силою еволюції.
Спостереження в природних умовах підтвердили адаптаційну реакцію популяцій на зміни умов довкілля. Так, у середині XIX сторіччя в Англії переважали особини метелика березового п’ядуна, що мали світле захисне забарвлення відповідно до тла стовбурів, на яких вони сидять. У промислових районах унаслідок задимлення стовбури беріз стали сірими, і за кілька десятків років у популяціях п’ядуна почала переважати форма з темними крилами (мал. 37.8). Завдяки ефективним заходам щодо зменшення промислових викидів в атмосферу берези в Англії знову посвітлішали, і в популяціях метелика тепер переважають світлокрилі особини.

Мал. 37.7. Внутрішньовидова мінливість черепашок суходільного кубинського слимака поліміти

Мал. 37.8. Темна та світла форми метелика березового п’ядуна на чистому(1) і задимленому (2) стовбурах беріз
Це свідчить про те, що з часом генофонд популяції змінився; в ньому знову поширився алель, який визначає світле забарвлення.
Які проблеми не змогла вирішити синтетична гіпотеза еволюції?
Синтетична гіпотеза не змогла пояснити, що собою становлять популяції прокаріотів та тих еукаріотів, яким не властива комбінативна мінливість, бо статевий процес або відсутній (наприклад, амеби, хлорела тощо), або відбувається самозапліднення (багато видів квіткових рослин) чи партеногенез. Тому механізми еволюції таких організмів до останнього часу практично не розглядали.

Мал. 37.9. Декілька підвидів тигра: 1 - балійський тигр. Цей підвид тигра мешкав на о-ві Балі (Індонезія). Останню самку застрелено у 1937 р.; 2 - яванський тигр. Мешкав на густонаселеному індонезійському о-ві Ява до початку 90-х років XX століття; кілька десятків особин збережено в зоопарках; 3 - каспійський (туранський) тигр. Мешкав виключно в прируслових лісах (тугаях). Останніх особин винищено наприкінці 50-х років XX століття. У зоопарках цей підвид відсутній
Отже, на характер еволюційних змін у популяціях впливають хвилі життя, дрейф генів та ізоляція, здатні змінювати генофонди популяцій. Якщо всі популяції обмінюються генетичною інформацією шляхом парувань особин з різних популяцій, то їхні генофонди будуть більш-менш подібні. А сам вид еволюціонуватиме як одне ціле. Якщо окремі популяції або їхні групи протягом багатьох поколінь відрізняються одна від одної за деталями будови, трофічними зв’язками, особливостями міграцій, термінами розмноження тощо, то їх часто згруповують у підвиди. Ми вже згадували, що існування підвидів - свідчення екологічної пластичності виду, тобто його здатності пристосовуватись до різноманітних умов довкілля (мал. 37.9).
Нові терміни та поняття. Синтетична гіпотеза еволюції, дрейф генів, ізоляція, мікроеволюція.
Запитання для повторення: 1. Які основні положення синтетичної гіпотези еволюції? 2. Чому популяцію вважають елементарною одиницею еволюції? 3. Що таке хвилі життя та дрейф генів? Яке їхнє значення в еволюції видів? 4. Що таке ізоляція та які її види? 5. Чому наявність підвидів вважають свідченням екологічної пластичності видів?
Проблемне завдання. Поміркуйте, чому наявність популяцій є необхідною умовою дивергенції виду.
Коментарі (0)